Выпуск 261
Лаборатория Наномир
Когда реальность открывает тайны,
уходят в тень и
меркнут чудеса ...
Была ли у Земли вторая Луна?

"День и ночь на этой планете перестали сменять друг друга 27 000
лунных месяцев после падения Луны." (Из фантастического рассказа "Заповедник под голубым солнцем")
Учитывая,
что Луна и Земли вращаются вокруг общего центра масс, нетрудно понять,
что в случае падения Луны на Землю центр масс продолжит двигаться по
орбите вокруг центра масс Солнечной системы.
Зная это, вернёмся к гипотезе астероидной зимы, в результате которой на Земле вымерло около половины биологических видов.
Мало
вероятно, что астероид упал на Землю, не изменив её скорости и
направления движения в Солнечной системе. Отсюда напрашивается вывод,
что это был не астероид, а вторая Луна. Её падение не могло изменить
параметров движения центра масс Земной системы.
Таким образом,
гибель динозавров и вместе с ними других биологических видов связана
именно с падением второй Луны. Соответственно, после её падения
наступила не астероидная, а "лунная" зима ...
Это
падение привело к увеличению радиуса Земли и расхождению материков.
После этого или предыдущего падения очередной луны на разных материках выжившие биологические виды оказались в
изоляции от других материков.
Обсудить на форуме.
Быстро ли падают луны?

Представим себе, что Луна уже начала падать на Землю. Быстро ли она упадёт?
Это
не праздный вопрос. Ведь с уменьшением расстояния до центра масс
орбитальная скорость будет возрастать, а это значит, что процесс падения
может затянуться. И чем меньше разность масс космических тел, тем
дольше будет продолжаться процесс падения.
Возможно, что в этом
процессе приливные силы полностью остановят собственное вращение
элементов двойной планеты. За счёт чего будет продолжаться падение? Как
будет диссипировать энергия вращения? Возможно, что этот процесс можно
смоделировать численными методами.
Представьте себе, что Луна
затормозила вращение Земли и постоянно находится в одной точке
небосвода. При этом постепенно падает на Землю. Сколько времени может
длиться это падение?
Многие знают, что падение космических станций
происходит достаточно быстро. Но это связано с их торможением в
атмосфере Земли. Если же Луна будет падать, оставаясь постоянно над
одной и той же точкой экватора Земли, то тормозить её будет нечем, и
падение может сильно затянуться. Конечно, Земля может начать
перетягивать вещество Луны, постепенно "высасывая Луну". При каком
расстоянии между центрами планет это начнётся?
Нетрудно
догадаться, что перетягивание вещества Луны начнётся в том случае, если
точка невесомости опустится ниже поверхности Луны. Попробуем рассчитать
эту точку для разных соотношений масс.
Если массы одинаковы, то
точка невесомости будет всегда находиться между планетами, т.е. даже
если они практически соприкоснутся. Но даже в этом случае вещество
планеты будет вспучиваться, т.к. под Луной вес грунта уменьшится.
При
сближении Луны и Земли на их поверхностях будут расти "приливные
холмы". Сегодня высота приливного холма в земном океане достигает
нескольких метров. Приливный холм на Луне в 20 раз выше. Интересно было
бы смоделировать процесс слияния двойной планеты и посмотреть, как долго
он будет длиться в случае разных соотношений масс.
Обсудить на форуме.
Продолжаем подготовку эксперимента в Дубне.

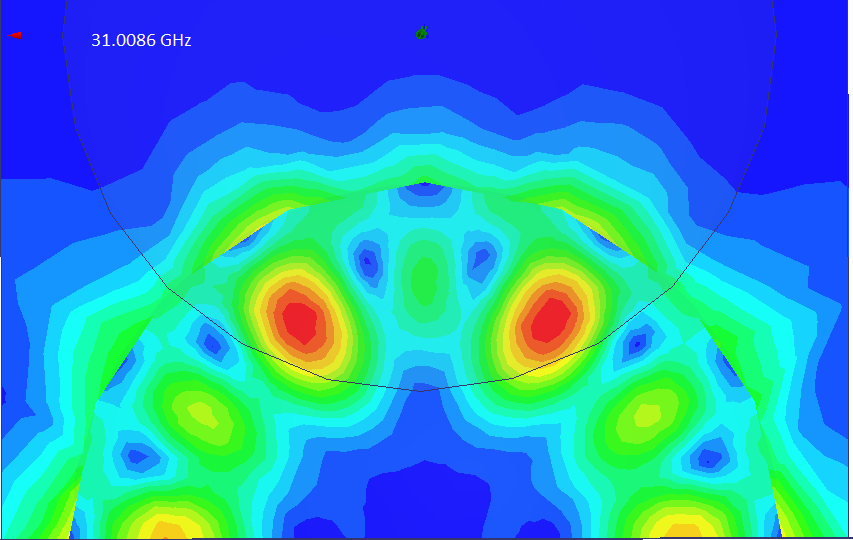
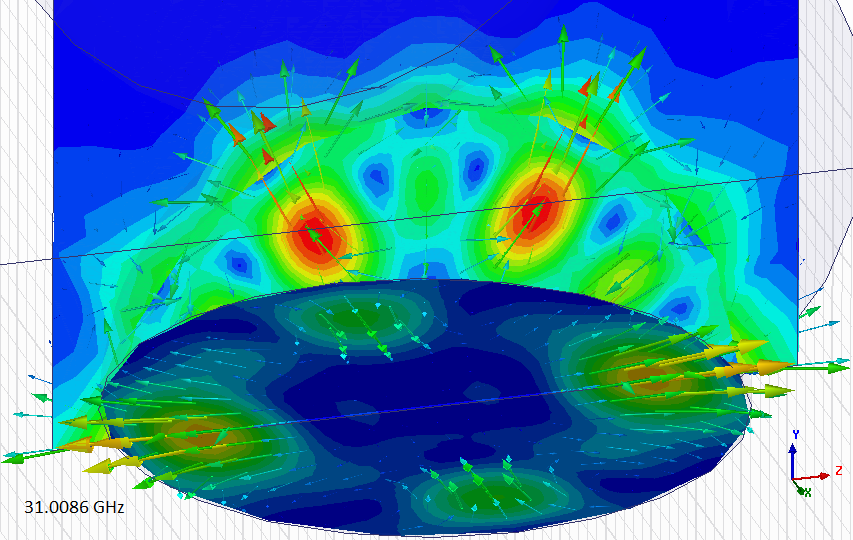
Колебательная мода 6HE в модели рубинового генератора обнаружена!

Программа
HFSS правильно показала распределение полей только в одном из 4 сферических резонаторов, но для наших целей этого достаточно.

Вспомогательные плоскости мешают правильной работе программы, но распределение поля искажается несильно.

Расчётная частота отличается от экспериментальной на ~3%

Распределение электрического
поля в двух ортогональных плоскостях (скалярное представление). Проверим, правильно ли направлены вектора электрического поля:

Да! Наконец, долгожданная колебательная мода 6HE найдена! Можно изучать распределение полей...

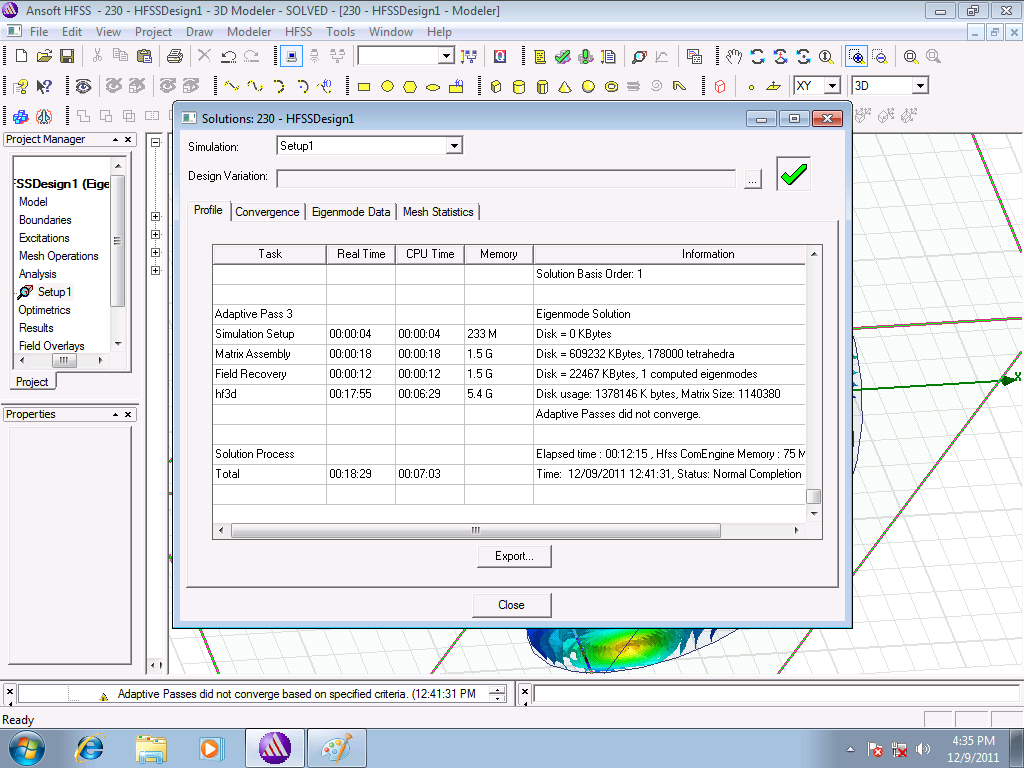
Я забыл добавить в модель диагональную вспомогательную плоскость.

Придётся
дополнить проект и повторить расчёт. К сожалению, вспомогательные
плоскости меняют расчётную сетку, что приводит к небольшому изменению
расчётной частоты. В таком случае можно заодно и отверстия в шариках
смоделировать. После настройки Windows расчёты ускорились в несколько
раз...
При задействованной оперативной памяти 5.4Gb расчётное
время уменьшилось со ~60 до 12 минут.
Обсудить на форуме.
Обсуждаем альтернативные модели тРНК.


Виктория: Я благодарю Вас за рассылку, если бы у меня был собственный журнал, я бы
тоже публиковала Ваши модели. А в чужих научных журналах шансов на
публикацию нет, рецензенты не пропустят даже как поисковое
моделирование.
Не хочу Вас расстраивать, но могу предложить только
соавторство в своих статьях. Только не обессудьте, буду писать, что
думаю и давать тот материал, который считаю проходным. Фантазии с
гиперзвуком рибосомы без экспериментального подтверждения никто даже
рассматривать не станет. По собственному опыту, даже на хорошо
обоснованный материал с достоверной статобработкой я получала
высмеивающие рецензии, что такого не может быть, потому что рецензент
(старикашка прошлого столетия) просто не в состоянии врубиться. У него и
раньше хитрости хватало только чтобы вскарабкаться по карьерной
лестнице и пригреться в тихом омуте, а теперь и подавно дементирующий
процесс поставил жирную точку на когнитивных способностях. Вердикт
такой: вот когда это опубликуют в зарубежном журнале, тогда мы
рассмотрим Вашу статью. К сожалению, это правило, а не исключение.
Вы
же ученый и сами понимаете, что в науке большинством голосов истинность
гипотез не выбирается. Обычно это делает эксперимент и время.
Кушелев: Соавторство тоже хорошо. Особенно, если сможете использовать модели, которые считаете подходящими для публикации.
Что касается "фантазии с гиперзвуком рибосомы", то в науке десятилетиями и столетиями публикуются аналогичные гипотезы. Та же
L-образная форма тРНК-гипотеза. Затраты на ацилирование в виде 2 порций
энергии по ~30 кДж/моль - тоже гипотеза. И так, куда ни ткни...
Виктория: Александр Юрьевич, а чем Вы объясняете закручивание АСС-конца тРНК при
присоединении аминокислотного остатка? Или этот хвост у тРНК по-Вашему и
без аминокислоты свёрнут в кольцо как у Шарика  ?
?

Кушелев: Я делал компьютерную модель по пластмассовой. А пластмассовую собирал
по интуиции. А интуиция подсказала, что аминокислоту держат два
цитозина. Я их рядом разместил, они, как оказалось, могут быть соединены
ковалентной связью. Тогда я сделал пикотехнологическую модель асс-конца
тРНК и стал думать, а куда же аденин присобачить? Стал его прикладывать
по-разному и вдруг понял, что его надо сзади к цитозину пристроить.
Кстати, позднее выяснилось, что это называется А-минорное
взаимодействие, которое я просто угадал в 1992-ом году. В 1992-ом году я
не заметил, что АСС-конец "загибается, как хвост у Шарика". Это стало
заметно на компьютерной модели, когда я стал складывать модель АСС-конца
из упрощённых моделей нуклеотидов. Ведь углы там строгие, поэтому я
стал поворачивать нуклеотиды, чтобы углы многогранных атомов сошлись.
Вот они и сошлись, но при этом АСС-конец "загнулся, как хвост у Шарика",
причём к оси симметрии тРНК. При этом атом азота с точностью до долей
ангстрема попал на ось вращения.
Может быть и в Вашей модели будет такое же "чудесное совпадение"? 

Виктория: В моей модели АСС-конца тРНК атом азота аланина тоже попадает на ось вращения акцепторного стебля.
Я могу прислать Вам этот файл и Вы сами в 3DS Max повращаете акцепторный
стебель вокруг его оси, проходящей через азот присоединённого аланина.
Вид вдоль оси симметрии акцепторного стебля. Если неправильно, то исправьте, пожалуйста.
Скачать файл для 3DS Max: 003.max
Виктория: Спасибо, очень эффектно. А.Ю., а можно сделать ролик, где было бы видно
вращение всей структуры из присланного мной файла (акцепторный стебель с
аминокислотой на АСС-конце + псевдоуридиловый стебель с псевдоуридиловой
петлёй) вокруг оси, проходящей через азот аланина и центр
псевдоуридиловой петли. Но! вращение должно быть не плавное, а медленное
и дискретное: три поворота на 120 градусов.
Заранее благодарна.
Кушелев: В Вашей модели аминокислота и атом азота не попадают
на ось симметрии акцепторного стебля. В этом случае аминокислота не
вставится в растущую белковую цепь. Пройдёт мимо Как быть?
Виктория: Это только для Вашей модели крестообразной тРНК актуально, чтобы ось
симметрии проходила через акцепторный стебель. В моей L-образной модели
тРНК достаточно, чтобы азот аланина принадлежал оси, соединяющей его с
условным центром псевдоуридиловой петли. Поскольку псевдоуридиловая
петля не является идеальной окружностью, то данная прямая не может
совпадать с осью симметрии самого акцепторного стебля, а немного
отклонена от неё, что не помешает аланину образовать пептидную связь в
рибосоме.
Кушелев: Как же не помешает, если поворот акцепторного стебля приводит к смещению
группы азота на несколько атомных диаметров. Атом азота не дотянется до
группы кислорода. Это всё равно, что ключ в замок вставлять с ошибкой
на 10 ключей по ширине. Никак не получится...
Виктория: Я же поясняла, что ось вращения НЕ обязана совпадать с осью симметрии
акцепторного стебля в моей модели тРНК, в отличие от Вашей. Проведите
другую ось вращения через атом азота и условный центр псевдоуридиловой
петли и посмотрите, что получится. Атом азота никуда не будет
смещаться. Он будет принадлежать оси вращения.
Кушелев: У Вас же вращение на 120 градусов происходит вокруг оси симметрии
акцепторного стебля, верно? А в этом случае атом азота должен лежать
именно на оси симметрии акцепторного стебля. Вы можете любые оси
проводить, но при повороте акцепторного стебля атом азота будет
вращаться именно вокруг оси симметрии акцепторного стебля. а значит не
попадёт на группу кислорода в двух случаях из трёх. Не верите? Проверьте
Виктория: НЕТ. А.Ю. это в Вашей модели ось симметрии акцепторного стебля и его ось
вращения совпадают, а в моей они НЕ могут совпадать. В моей модели тРНК
поворот акцепторного стебля реализуется не индивидуально, а вследствие
поворота псевдоуридиловой петли относительно дигидроуридиловой.
Центральная точка взаимодействия этих петель в L-образной структуре тРНК
НЕ принадлежит оси симметрии акцепторного стебля. Поэтому и ось
вращения последнего НЕ совпадает с осью симметрии, а проходит через выше
означенную точку и азот аланина. Поэтому поворот по такой оси вращения
НИКУДА НЕ смещает азот аланина, который сам лежит на этой оси. Это ведь
очевидно. Вы со мной согласны?
Кушелев: А Вы покажите, как конкретно поворачивается в Вашей модели акцепторный
стебель тРНК. Кстати, даже если Вы будете поворачивать его прямо вокруг
центра атома азота, то это не спасёт Вашу модель. Дело в том, что атом
азота у Вас сам неправильно ориентирован. Ведь его 8-электронная
оболочка должна совмещаться сама с собой при повороте на 120 градусов.
Она не совмещается. Ни при вращении вокруг оси симметрии акцепторного
стебля, ни вокруг оси, соединяющий его центр с точкой соединения
акцепторного и антикодонового стеблей. Так что Вам нужно не только
объяснить вращение акцепторного стебля вокруг не его оси симметрии, но и
что-то делать с ориентацией атома азота...
Уважаемая
Виктория! Обратите внимание, как ориентирован атом азота в моей модели и
в Вашей. Он должен вращаться вокруг оси симметрии третьего порядка. А у
Вас? Чтобы всё это исправить, "костылей не хватит"
Файл-сцена для 3DS Max: 003.max
И
это ещё не все проблемы, которые есть в Вашей модели. При движении тРНК
в физрастворе аминокислота должна быть защищена. Если она находится на
оси вращения тРНК и прикрыта сверху фосфатной группой, с которой её
соединяет водородная связь, как в моей модели, то вероятность "доехать"
до рибосомы у неё будет несоизмеримо выше, чем если она будет
незащищённой и двигаться по замысловатой траектории. Её просто "сдует" с
АСС-конца тРНКВиктория:
Это всё только потому, что Вы продолжаете вращать мою модель
вокруг оси симметрии акцепторного стебля. Попробуйте ориентировать
аланин в моей модели аналогично тому, как у Вас, но только в составе
моей модели ,
и вращать по его оси третьего порядка. Увидите, как будет вращаться при
этом акцепторный стебель. Мне кажется, что псевдоуридиловая петля на
противоположном конце моей модели при этом должна описывать окружность, а
не просто вращаться. Вращение акцепторного стебля с аминокислотой на АСС-конце в моей модели
можно представить в виде вращающегося лассо: аминокислота в точке
удержания верёвки (в кулаке), а сама петля лассо -- это псевдоуридиловая
петля. Вращение петли (и аминокислоты) происходит не вокруг оси
симметрии, принадлежащей верёвке, их соединяющей, а вокруг оси вращения.
Это надо нарисовать.
Кушелев: Вы наклоните модель тРНК как
считаете нужным и посмотрим, как она будет вращаться вокруг оси третьего
порядка атома азота. Мне тоже очень интересно. Кстати, у атома азота 4
оси третьего порядка. Вам надо выбрать, вокруг какой из них вращать
модель тРНК...
Виктория: На счет "сдувания" аминокислоты с хвоста
тРНК, то это фантазия. Броуновское движение не способно разрушить ни
одной ковалентной связи в составе молекул любой сложности.
Кушелев:
А мы с Вами уже рассчитали, что кинетической энергии тРНК с
десятикратным запасом хватает, чтобы разорвать ковалентную связь и
отрезать триплет иРНК. Это значит, что достаточно зацепиться
аминокислотой за другую макромолекулу, например, тоже тРНК, и
аминокислота оторвётся, не доехав до рибосомы...
Виктория: А.Ю., мы действительно с Вами это обсуждали ранее, но Вы, к сожалению,
остались при своих фантазиях и расчетах, которые опираются на
игнорировании объективных экспериментальных данных о вязкости
цитоплазмы, третичной структуре тРНК и прочности ковалентных связей.
Кушелев: Какие проблемы? Я пользовался экспериментальными данными о
скоростях движения молекул при комнатной температуре. Если у Вас есть
другие данные и другой расчёт, давайте сравним результаты.
Виктория: А можно выбрать ту же ось третьего порядка, что и у Вас, чтобы сравнивать подобное с подобным.
Кушелев: Вот, что получится, если вращать Вашу модель вокруг ближайшей оси симметрии третьего порядка атома азота: 006.max
Виктория: Первая подсказка -- три варианта поворота акцепторного стебля с аминокислотой (Вы смоделировали в сообщении 315)
Кушелев: Так акцепторный
стебель поворачиваться не может. Он же оторвётся от
антикодонового. Противоположный аминокислоте конец акцепторного стебля
должен поворачиваться вокруг собственной оси симметрии.
Виктория: При функционировании таких структурно специфичных тРНК наблюдается
простота и единообразие без вращений и обрезаний, что обеспечивает
высокую скорость и точность трансляции.
Кушелев: О какой скорости Вы говорите, если непонятно даже, как двигается по
Вашей модели тРНК? Ведь при броуновском движении она может никогда не
добраться до рибосомы. Другое дело - поступательное движение винта с
управлением гиперзвуковым сигналом рибосомы.
Виктория: Разумеется в одной тРНК поворот акцепторного стебля реализуется при
формировании её третичной структуры и далее не меняется. Но, в другом
варианте изоакцепторной тРНК для той же аминокислоты в структуре
изначально реализован другой поворот акцепторного стебля за счет
поворота псевдоуридиловой петли относительно дигидроуридиловой по
часовой стрелке и фиксации угла поворота уже другими межпетельными
комплементарными парами, который также не меняется в дальнейшем, . При
этом ничего ниоткуда не отрывается.
Кушелев: Попробуйте сделать модели для трёх вариантов композиционного кода.
Очень интересно, как будут выглядеть модели трёх разных тРНК.
Если начало акцепторного стебля, на конце которого находится
аминокислота, будет у всех трёх тРНК в одном месте прикреплено к
антикодоновой петле, то ориентация атома азота будет неправильной. Это
Вы понимаете?
Если же Вы будете прикреплять начало акцепторного
стебля к антикодоновому в том месте, где он окажется, поворачиваясь
вокруг оси третьего порядка атома азота, то начало просто не дотянется в
двух случаях из трёх, т.е. радиус окружности, по которой поворачивается
начало акцепторного стебля в несколько раз превышает толщину стеблей
тРНК.
"Тут всю систему менять надо..."
Виктория: Акцепторный и антикодоновый стебли НЕ лежат на одной прямой и НЕ
формируют единого двуспирального луча. Они максимально удалены друг от
друга и располагаются под углом около 90 градусов. Вся остальная часть
тРНК как раз находиться в узле между ними.
Кушелев:
Представьте себе, что тРНК это у нас балерина.
При
повороте ноги вокруг оси симметрии угол наклона большого пальца не
совпадает с осью вращения, как в Вашей модели тРНК. Что произойдёт с
ногой, если её вращать вокруг оси третьего порядка, проходящей вдоль
кости большого пальца ноги? Правильно. Оторвётся нога в тазобедренном
суставе.

Виктория: А.Ю., "две ноги" тРНК соединяются между собой "тазобедренным суставом"
из псевдоуридиловой и дигидроуридиловой петель, который не лежит в одной
плоскости с лучами, а ориентирован под углом к ним. Поэтому поворот
акцепторного стебля с аминокислотой в составе одного из лучей происходит
при вращении одной плоскости "сустава" (псевдоуридиловой петли)
относительно другой (дигидроуридиловой петли) по часовой стрелке.
На
Вашей картинке с балериной, эта девочка может вполне сделать и ласточку,
и вертикальный шпагат, при которых нога может повернуться даже пяткой
вперёд и НЕ оторваться .
Кушелев: Но для этого ногу надо поворачивать вокруг оси, проходящей через
тазобедренный сустав. Если же её поворачивать вокруг оси симметрии
указательного пальца ноги (аналог оси третьего порядка атома азота), то
нога оторвётся:
Закон геометрии однако...
Обсудить на форуме.
Кто профинансирует включение гранатового генератора в микроволновой
печи?
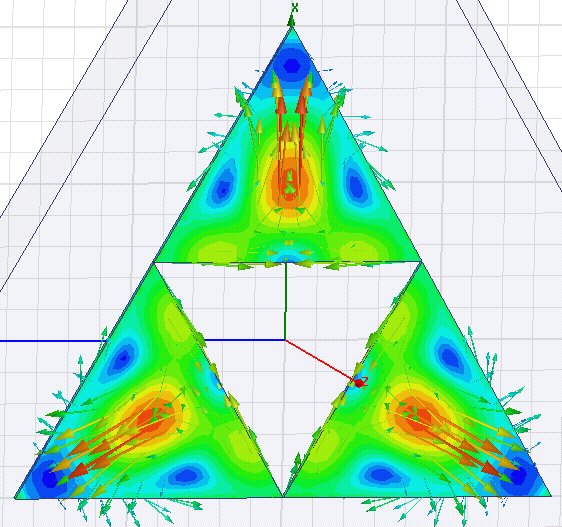
Модель резонатора "Гранатовый треугольник". Колебательная мода "Саваофа"

Можно
ли изготовить элементы гранатового резонатора на частоту 2.45 ГГц из
куска иттрий-алюминиевого граната ИАГ-Ce (жёлтого цвета) ? Как показали
расчёты в программе HFSS, габариты элементов треугольной формы должны
быть не менее 100*70 мм. В данную заготовку они не вписываются. Придётся
покупать сырьё более крупных размеров массой ~1.5 кг.

Фирма Meller Optics производит круглые пластины из сапфира с максимальным диаметром 76.2 мм.
Это почти тот размер, который нужен. Но для гранатового генератора
прозрачный лейкосапфир не подходит. Нужен сильно окрашенный
монокристалл, например, рубин, шпинель, гранат. Наиболее доступным по
цене кристаллом подходящего размера является гранат, выращенный по
методу Багдасарова.
Кто из соинвесторов оплатит
кристалл-заготовку, тот будет иметь непосредственное отношение к результатам эксперимента на частоте 2.45 ГГц :)
Лаборатория
Наномир объявляет тендер на покупку сырья для эксперимента на частоте
2.45 ГГц. У соинвесторов есть возможность объединить усилия, т.е.
совместно профинансировать этот эксперимент.
Приглашение к сотрудничеству
для людей умеющих самостоятельно мыслить; не просто умных, а мудрых, которые чувствуют, где истина
Лаборатория
Наномир готова к любому взаимовыгодному сотрудничеству. У нас
есть сторонники как явные, которые помогают морально и
материально, есть очень много пассивных наблюдателей, есть и ярые
противники, которые используют
любые методы и средства (аморальные и просто преступные), чтобы
уничтожить работу лаборатории и дискредитировать ее.
В одиночку
внедрить технологии, выводящие цивилизацию на новый уровень,
невозможно. Благодаря поддержке множества заинтересованных людей
проделана огромная работа. Ознакомиться с её результатами можно изучив
материал рассылки "Новости лаборатории Наномир". Люди науки могут изучить научные
труды.
Вклад каждого не останется незамеченным в
случае успеха в реализации научных проектов. Результаты совместной
деятельности принадлежат участникам проекта пропорционально
коэффициентам творческого и финансового участия.
В этом году были
куплены рубиновые шарики для эксперимента на сумму ~1000 долл. В
результате было сделано научное открытие, проверена защита
диэлектрических резонаторов от перенапряжения. В этом же году, вероятно,
можно будет создать микроволновую энергетику,
т.к. удалось найти сырьё (рубин #8), из которого сделаны рубиновые
шарики для эксперимента в Дубне.
28 сентября начался эксперимент
по созданию "эликсира вечной молодости". Благодаря первому взносу (в
размере 500 долларов) Золдракса и поддержке других соинвесторов.
Продолжаются переговоры с потенциальными инвесторами по поводу
финансирования этого проекта.
Созданы первые версии пикотехнологии, с помощью которой Александр Кушелев и Виктория Соколик сделали более10 научных открытий.
Сотрудничество
может быть различным:
- участие в научных дискуссиях на форуме (конструктивное)
- совместное создание коммерческого продукта
- поиск инвесторов
- выступить менеджером по продаже готовых коммерческих продуктов
- конструктивные предложения по продвижению идей лаборатории Наномир
- содействие в проведении экспериментов и т.п.
- написание совместных научных статей и т.п.
- материальный вклад (денежный или обеспечение оборудованием и материалами)
Пожалуйста, сообщайте о своем вкладе, чтобы мы зачли Вас как партнера лаборатории Наномир.
+7-926-5101703, +7-903-2003424, +7-916-8265031, Skype: Kushelev2009, mail: kushelev2011@yandex.ru
веб-мани: WM-кошелек R426964799301